Estrutura e funções dos tecidos vegetais
Um violento temporal, uma seca prolongada, um animal herbívoro ou qualquer outro agente agressivo do meio, têm que ser enfrentados pela planta imóvel, ao contrário de um animal, que pode se refugiar em lugar seguro até que as condições ambientais se normalizem.
Os tecidos protetores, ou de revestimento, de uma traqueófita são a epiderme e o súber. A eficiência deles pode garantir a proteção da planta contra diversos agentes agressivos do meio.
Os tecidos protetores, ou de revestimento, de uma traqueófita são a epiderme e o súber. A eficiência deles pode garantir a proteção da planta contra diversos agentes agressivos do meio.
O súber
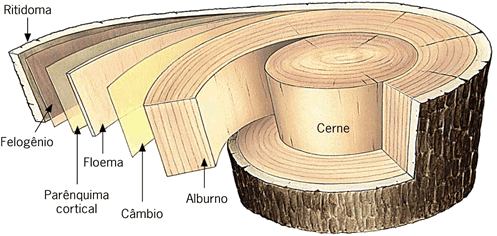
O súber é um tecido secundário, muito leve e elástico, formado pelo câmbio suberofelogénico e apenas presente em caules lenhosos.
As células do súber são mortas devido à deposição na parede secundária de suberina. A suberina é uma substância lipídica, tornando estas células impermeáveis aos gases e à água.
Ao contrário da epiderme, o súber é um tecido com diversas camadas de células, podendo atingir espessuras importantes, como no caso dos carvalhos ou dos sobreiros, onde forma a cortiça.
Quando se forma, o súber substitui a epiderme nas suas funções de protecção, impedindo a perda de água e protegendo o frágil floema.
Dado que se trata de um tecido impermeável, é necessário que estas camadas de células sejam interrompidas a espaços regulares, possibilitando as trocas gasosas com o meio. Essas zonas de interrupção designam-se lentículas

O tronco de uma árvore periodicamente cresce em espessura. Esse crescimento força a ruptura do súber que racha em muitos pontos e acaba se destacando, juntamente com outros tecidos. Antes, porém, a árvore elabora novo súber que substituirá o que vai cair. A este material periodicamente destacado dá-se o nome de ritidoma.
A epiderme
A epiderme das plantas vasculares é um tecido formado, de modo geral, por uma única camada de células de formato irregular, achatadas, vivas e aclorofiladas. É um tecido de revestimento típico de órgãos jovens (raiz, caule e folhas). A epiderme de uma raiz mostra uma camada cilíndrica de revestimento, com uma zona pilífera, cujos pelos nada mais são do que extensões de uma célula epidérmica.
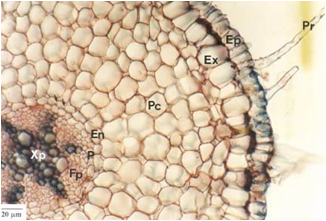
Corte transversal da raiz primária de Mandevilla velutina. Ep = epiderme; Pr = pêlos radicular; Ex = exoderme; Pc = parênquima cortical; En = endoderme; P = periciclo; Xp = xilema primária; Fp = floema primário.
Caules jovens também são revestidos por uma fina epiderme não-dotada, porém, de pelos.
É na folha que a epiderme possui notáveis especializações: sendo um órgão de face dupla, possui duas epidermes, a superior e a inferior.
É na folha que a epiderme possui notáveis especializações: sendo um órgão de face dupla, possui duas epidermes, a superior e a inferior.
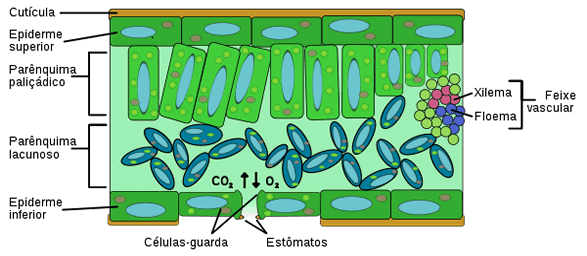
As células epidérmicas secretam para o exterior substâncias impermeabilizantes, que formam uma película de revestimento denominada cutícula. O principal componente da cutícula é a cutina, um polímero feito de moléculas de ácidos graxos. Além de evitar a perda de água, a cutícula protege a planta contra infecções e traumas mecânicos.
Diferenciam-se na epiderme estruturas como estômatos, tricomas, hidatódios e acúleos.
Estômatos

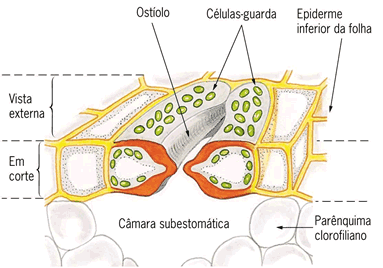
Estômato com células-guarda permitindo a abertura do ostíolo
O primeiro tecido de revestimento das plantas é a epiderme, formada por uma única camada de células diretamente ligadas umas às outras. A folha de uma planta tem dois lados diferentes, o lado de cima - a epiderme - é revestido por uma cutícula que a protege da perda de água, por isso é lisinha e brilhante como se estivesse encerada; a parte de baixo apresenta uma estrutura responsável por realizar as trocas gasosas com o ambiente, é conhecida como estômato.
O estômato é constituído por duas células com formato parecido com o do feijão, meio curvadinho, essas células são chamadas de células-guarda; que são ricas em clorofila e estão inseridas na epiderme junto com inúmeras outras células conhecidas como células acessórias, que não possuem clorofila.
O orifício formado pelo curvamento das células-guarda dos estômatos é conhecido como ostíolo e é por essa abertura que ocorrem as trocas gasosas entre os gases presentes na atmosfera e aqueles contidos no interior da planta.
A abertura dos estômatos depende de diversos fatores, principalmente da luminosidade, concentração de gás carbônico (CO2) e da disponibilidade de água nas raízes. A maioria das plantas abre seus estômatos pela manhã e os fecha com a diminuição da luminosidade, permitindo a absorção do gás carbônico durante o dia para a realização da fotossíntese e reduzindo a perda de água para o ambiente durante a noite.
A quantidade de água disponível nas raízes é um fator determinante para a manutenção do equilíbrio hídrico das plantas, caso não haja água em quantidade suficiente, os estômatos são fechados para impedir a transpiração, mesmo que haja luz para a realização da fotossíntese. Isso explica o fato de algumas plantas perderem suas folhas durante períodos de seca ou escassez de água.
Em relação ao gás carbônico, quando esse começa a se acumular dentro da folha é um indicativo de que a taxa de fotossíntese está diminuindo e que a abertura dos estômatos não é mais necessária. Assim, durante o dia, com a luz e a ocorrência da fotossíntese, há pouco CO2 no mesófilo foliar, então, estômato aberto; mas quando a situação se inverte, os estômatos se fecham.
Mas por que o estômato se abre?
O movimento das células-guarda se constitui no efeito causado pela entrada ou saída de saís de potássio nas células. Quando a concentração de potássio aumenta, as células-guarda absorvem água e ficam inchadas, modificando sua forma e permitindo a formação do ostíolo e a troca de gases com o meio externo. Quando a quantidade de potássio reduz a célula, ela cede água para as células acessórias e retornam à sua forma, fechando assim o ostíolo.
Atenção! A troca de gases entre a planta e o meio ocorre através dos estômatos da epiderme e de uma estrutura chamada lenticelas presentes no súber.
As lenticelas são pequenas aberturas que facilitam o ingresso e a saída de gases nas raízes e caules suberificados. |
Tricomas
Os tricomas são geralmente estruturas especializadas contra a perda de água por excesso de transpiração, ocorrendo em planta de clima quente. Podem ser, no entanto, secretores, produzindo secreções oleosas, digestivas ou urticantes. As plantas carnívoras possuem tricomas “digestivos” e a urtiga, planta que provoca irritação da pele, possui tricomas urticantes.

Acúleos
Os acúleos, estruturas pontiagudas com função de proteção da planta contra predadores, são frequentemente confundido com espinhos, que são folhas ou ramos modificados. Os acúleos são fáceis de destacar e são provenientes da epiderme. Podem ser encontrados nas roseiras.
Hidatódios
Hidatódios são estômatos modificados, especializados em eliminar excessos líquidos da planta. Os hidatódios geralmente presentes nas bordas das folhas, onde, pela manhã, é possível observar as gotas de líquido que eles eliminam, fenômeno conhecido como gutação.
A sustentação das traqueófitas
O porte das traqueófitas só foi possível por adaptações que tornaram possível a sustentação do organismo vivo e a disponibilidade e transporte de água para todas as células.
A sustentação de uma traqueófita é devida à existência de tecidos especializados para essa função: ocolênquima e o esclerênquima.
O Colênquima
O colênquima é derivado de um tecido parenquimatoso. Quando cortadas transversalmente podem ter aspectos variados. Como na maioria das células dos tecidos vegetais, as células do colênquima são vivas apenas na maturidade. Por serem vivas na maturidade podem continuar desenvolvendo paredes espessas, enquanto o órgão está se alongando, o que torna essas células adaptadas para a sustentação de órgãos jovens em crescimento.
 É comum este tecido ocorrer em feixes isolados contínuos ou então como “cilindros” abaixo da epiderme, nos caules e pecíolos (estrutura da folha que une o limbo ao caule), podendo também ser encontrados na margem das nervuras das folhas eudicotiledôneas. Suas células podem ter também a forma alongada, que auxilia na sua função que é de sustentação esquelética dos vegetais. Apresentam protoplastos vivos e podem possuir cloroplasto.
É comum este tecido ocorrer em feixes isolados contínuos ou então como “cilindros” abaixo da epiderme, nos caules e pecíolos (estrutura da folha que une o limbo ao caule), podendo também ser encontrados na margem das nervuras das folhas eudicotiledôneas. Suas células podem ter também a forma alongada, que auxilia na sua função que é de sustentação esquelética dos vegetais. Apresentam protoplastos vivos e podem possuir cloroplasto.
Podemos ter cinco tipos de colênquima que são:
- Colênquima anelar: Por toda a borda da célula um espessamento uniforme.
- Colênquima angular: Espessamento por todos os ângulos da célula.
- Colênquima lamelar: Nas bordas das células há maiores espessamentos tangenciais.
- Colênquima lacunar: espessamento nas paredes próximas ao espaço intercelular.
- Colênquima radial: células alongadas e paralelas, alocadas radialmente.

Colênquima angular
Alguns fatores como, regiões jovens, ventos fortes e como a herbivoria e infestação de organismos que prejudicam as folhas estimulam o espessamento das paredes. Regiões jovens, são fotossintetizantes, por isso necessitam de um tecido que possa haver a passagem de luz, e no caso o colênquima seria este tecido. As paredes do colênquima são altamente hidrofílicas (interagindo fortemente com a água).
As fibras colenquimáticas geralmente aparecem em corte transversal muito próximo a epiderme.
À medida que as células deste tecido vão envelhecendo seu espessamento podem ser alterados, e dependendo se as células forem bastante velhas podem até se transformarem em esclerênquima, pela destituição de paredes secundárias lignificadas.
O Esclerênquima

Na maturidade, a elasticidade torna-se mais importante que a plasticidade, pois assim a parede pode ser deformada por tensão ou pressão, reassumindo sua forma em condições normais.
Assim, vento, passagem de animais, etc., não causam deformações definitivas nas plantas.
Além dessa característica, o esclerênquima forma uma camada protetora ao redor do caule, sementes e frutos imaturos, evitando que animais e insetos se alimentem deles, pois a lignina não é facilmente digerida.
Ocorrem em faixas ou calotas ao redor dos tecidos vasculares e também em tecidos parenquimáticos, como na medula, caule e pecíolo de algumas plantas. Oferece sustentação e proteção.
CARACTERÍSTICAS
Células com protoplastos mortos na maturidade, parede secundária lignificada (a lignina é uma substância amorfa, formada pela polimerização de vários álcoois e confere maior rigidez à parede). A forma das células é muito variável.
TIPOS
1- Esclereídes ou esclerócitosCélulas curtas, espessadas, com numerosas pontoações. O tecido formado é muito rígido. A textura pétrea da pêra é devida à presença de inúmeros esclerócitos isodiamétricos na polpa.2- FibrasCélulas longas, com extremidades afiladas, lume reduzido e paredes secundárias espessas. Servem como de elemento de sustentação nas partes vegetais que não mais se alongam.Podem se originar do pró-câmbio, sendo chamadas fibras do floema ou xilema primários ou do câmbio, sendo denominadas fibras do xilema ou floema secundários; além disso, células do parênquima cortical, mesofilo e epiderme podem originar fibras.
Em Linum, por exemplo, as fibras se originam no floema e são a fonte do linho. As fibras do cânhamo (Cannabis sativa) se desenvolvem entre células de floema e também a partir do câmbio. Admite-se que, tanto nas fibras quanto nas esclereídes, após o completo desenvolvimento de suas paredes secundárias, o protoplasto, não mais funcional, seja eliminado.
Estrutura interna das folhas
A folha é totalmente revestida pela epiderme, e seu interior, denominado mesófilo (do grego, mesos, meio e phylon, folha), é constituído por parênquima clorofiliano, tecidos condutores e tecidos de sustentação.
O parênquima clorofiliano foliar pode ser, em geral de dois tipos:
O parênquima clorofiliano foliar pode ser, em geral de dois tipos:
- palisádico - constituído por células prismáticas e justapostos como uma paliçada, e
- lacunoso -constituído por células de forma irregular, que deixam espaços ou lacunas entre si.
Pode haver parênquima paliçádico junto à epiderme de ambas as faces da folha, ou, como é mais comum, parênquima paliçádico junto a epiderme da face superior e lacunoso junto à inferior.
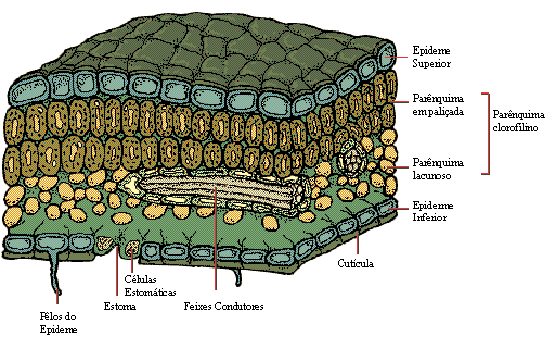
Nervuras foliares
Os tecidos condutores presentes na folha encontram-se agrupados em feixes libero-lenhosos, nos quais o xilema está voltado para a epiderme superior e o floema, para a epiderme inferior. Os feixes condutores mais grossos formam as nervuras foliares, visíveis a olho nu.
Os Tecidos Condutores de Água e de Nutrientes em Traqueófitas
Além das trocas gasosas, um dos maiores problemas de um vegetal terrestre relaciona-se à disponibilidade de água e sua perda, pois para à realização da fotossíntese é fundamental que se consiga, além do gás carbônico, a água. O problema de perda de água através das folhas é, em parte, minimizado pela presença de cutículas lipídicas, nas faces expostas das epidermes, que as impermeabilizam. Porém, isso dificulta as trocas gasosas.
A existência nas traqueófitas de aberturas epidérmicas reguláveis (os estômatos) que permitem as trocas gasosas e ao mesmo tempo ajudam a evitar perdas excessivas de vapor de água é um mecanismo adaptativo importante.
O transporte de água e nutrientes em uma traqueófita ocorre em parte por difusão de célula à célula e, na maior parte do trajeto, ocorre no interior de vasos condutores.
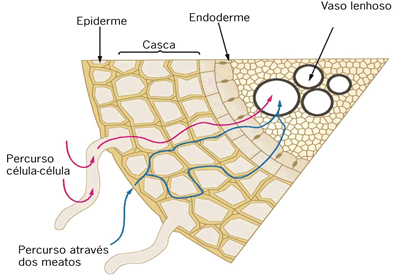
Inicialmente, ocorre a absorção de água e nutrientes minerais pela zona pilífera da raiz. Os diferentes tipos de íons são obtidos ativa ou passivamente e a água é absorvida por osmose.
Forma-se uma solução aquosa mineral, a seiva bruta ou seiva inorgânica. Essa solução caminha de célula a célula radicular até atingir os vasos do xilema (ou lenho) existentes no centro da raiz. A partir daí, o transporte dessa seiva ocorre integralmente dentro dos vasos lenhosos até as folhas. Lá chegando, os nutrientes e a água difundem-se até as células e são utilizados no processo da fotossíntese.
Os compostos orgânicos elaborados nas células do parênquima clorofiliano das folhas difundem-se para outro conjunto de vasos do tecido condutor chamado floema ou líber. No interior dos vasos liberianos, essa seiva orgânica ou seiva elaborada é conduzida até atingir as células do caule, de um fruto, de um broto em formação, de uma raiz etc., onde é utilizada ou armazenada.
O xilema
O xilema é um tecido responsável por conduzir água e nutrientes inorgânicos em plantas vascularizadas. Este tecido está expandido ao longo de todo o corpo vegetal, constituindo um sistema contínuo que armazena algumas substâncias e confere sustentação ao vegetal. Se considerarmos o corpo primário, resultante do crescimento primário da planta, pode-se dizer que o xilema nasce a partir do procâmbio (um meristema primário), porém se levarmos em conta o corpo secundário temos que este tecido condutor se originará a partir do câmbio vascular.

Já os elementos de vaso são perfurados e isso confere mais eficácia na hora de transportar a água ao longo da planta. Esses furos permitem um trânsito livre da água de um elemento de vaso a outro. Esses são pontos positivos, mas também há os pontos negativos dos elementos de vaso. Podemos exemplificar enfatizando um contraponto deste sistema tão aberto: a fragilidade. As traqueídes são mais estreitas e menores que os elementos de vaso, por isso somam mais pontos nesse quesito, trazem mais “tranquilidade” para a planta. O motivo de tanta preocupação é que durante o trânsito da água nesses vasos pode ocorrer a formação de pequenas bolhas de ar. Quando isto ocorre na traqueíde há a obstrução da mesma, porém como é pequena e normalmente curta não tem muita influência para o contexto do vegetal. Porém o problema está quando estas bolhas se formam nos elementos de vaso, pois obstruem o tráfego de água não só no ponto em que a bolha está, mas também por todo o vaso. Então esse vaso obstruído (largo e comprido) fará falta para a planta.
Anteriormente foi falado sobre a armazenagem de substâncias pelo xilema. Isto só é possível porque este tecido também conta com células parenquimáticas na sua constituição, são essas células que armazenam muitas substâncias num primeiro instante, mas depois quando amadurecem também desempenham um outro papel importante: a sustentação.
A condução da seiva inorgânica
Vimos que as raízes absorvem água do solo através da região dos pelos absorventes ou zona pilífera. Desta, a água atravessa as células do córtex, endoderme e periciclo da raiz. Na endoderme o fluxo da água pode ser facilitado ela existência das chamadas células de passagem. A água atinge os vasos do xilema e, a partir desses vasos, atinge a folha. Na folha, ou ela é usada na fotossíntese ou é liberada na transpiração.
Atribui-se a condução da seiva inorgânica (ou bruta) a alguns mecanismos: pressão da raiz, sucção exercida pelas folhas e capilaridade.
- A pressão da raiz – O movimento da água através da raiz é considerado como resultante de um mecanismo osmótico. A água que está no solo entra na célula do pêlo radicular, cuja concentração é maior que a da solução do solo. A célula radicular é menos concentrada que a célula cortical. Esta, por sua vez, é menos concentrada que a célula endodérmica e, assim por diante, até chegar ao vaso do xilema, cuja solução aquosa é mais concentrada de todas nesse nível. Assim, é como se a água fosse osmoticamente bombeada, até atingir os vasos do xilema.
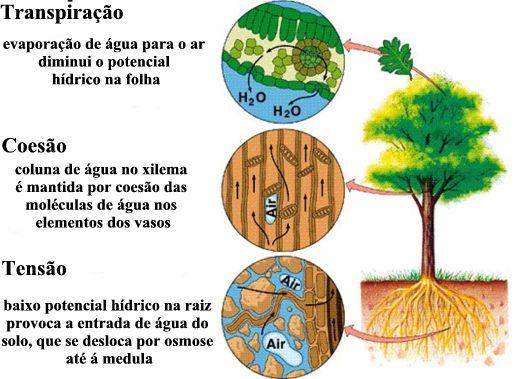
- A sucção exercida pelas folhas – A hipótese mais aceita, atualmente, para o deslocamento da seiva do xilema é baseada na “sucção” de água que a copa exerce. Esta “sucção” está relacionada com os processos de transpiração e fotossíntese que ocorrem nas folhas. Para que essa “aspiração” seja eficiente, dois pré-requisitos são fundamentais: inexistência de ar nos vasos de xilema e uma força de coesão entre as moléculas de água. A coesão entre as moléculas de água faz com que elas permaneçam unidas umas às outras e suportem forças extraordinárias, como o próprio peso da coluna líquida no interior dos vasos, que poderiam levá-las a separar-se. A existência de ar nos vasos do xilema romperia essa união e levaria à formação de bolhas que impediriam a ascensão da seiva lenhosa. As paredes dos vasos lenhosos igualmente atraem as moléculas de água e essa adesão, juntamente com a coesão, são fatores fundamentais na manutenção de uma nova coluna contínua de água no interior do vaso.
- A transpiração e a fotossíntese removem constantemente água da planta. Essa extração gera uma tensão entre as moléculas de água já que a coesão entre elas impede que se separem. A parede do vaso também é tracionada devido à adesão existente entre ela e as moléculas de água. Para que se mantenha a continuidade da coluna líquida, a reposição das moléculas de água retiradas da copa deve ser feita pela raiz, que, assim, abastece constantemente o xilema.
- O efeito da capilaridade na condução da seiva – Os vasos lenhosos são muito delgados, possuem diâmetro capilar. Assim, a ascensão do xilema ocorre, em parte, por capilaridade. No entanto, por esse mecanismo, a água atinge alturas bem inferiores a 1 metro e, isoladamente, esse fato é insuficiente para explicar a subida da seiva inorgânica.
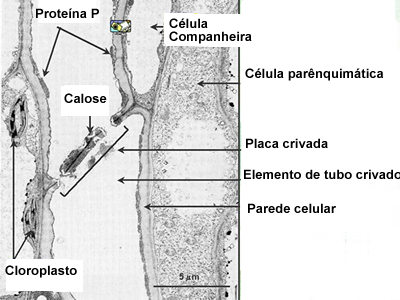
O floema
Floema é o principal tecido condutor de substâncias orgânicas das plantas vasculares. Como o xilema, o floema também pode ser primário ou secundário e suas principais células (que são originadas do meristema apical) responsáveis pela condução da seiva são os elementos crivados. Sendo que esses elementos podem ainda ser de dois tipos: células crivadas ou elementos do tubo crivado. Curiosamente recebem essa denominação pela semelhança que o aglomerado de poros em sua estrutura tem com um “crivo”. É por esses poros que se faz possível conectar protoplastos de células vizinhas (ou companheiras, como na figura ao lado). Algumas denominações (nomenclaturas) serão detalhadas a seguir: Sendo que esses elementos podem ainda ser de dois tipos: células crivadas ou elementos do tubo crivado. Curiosamente recebem essa denominação pela semelhança que o aglomerado de poros em sua estrutura tem com um “crivo”. É por esses poros que se faz possível conectar protoplastos de células vizinhas (ou companheiras, como na figura ao lado). Algumas denominações (nomenclaturas) serão detalhadas a seguir:

O floema pode ser facilmente identificado, por exemplo, nos vegetais que apresentam um crescimento lateralizado (ou secundário) já que é a própria casca que recobre o caule desta planta.
|
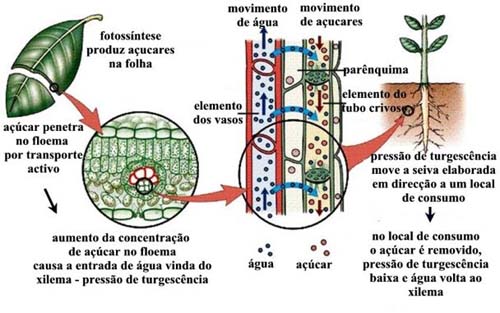
A condução da seiva elaborada
A seiva orgânica, elaborada no parênquima das folhas, é lançada nos tubos crivados do floema e conduzida a todas as partes da planta que não são auto-suficientes. O transporte é orientado principalmente para a raiz, podendo haver algum movimento em direção ao ápice do caule e folhas em desenvolvimento. De modo geral, os materiais orgânicos são translocados para órgãos consumidores e de reserva, podendo haver inversão do movimento (isto é, dos órgãos de reserva para regiões em crescimento), quando necessário.
A hipótese de Münch
A hipótese mais aceita atualmente para a condução da seiva elaborada é a que foi formulada por Münch e se baseia na movimentação de toda a solução do floema, incluindo água e solutos. É a hipótese do arrastamento mecânico da solução, também chamada de hipótese do fluxo em massa da solução. Por essa hipótese, o transporte de compostos orgânicos seria devido a um deslocamento rápido de moléculas de água que arrastariam, no seu movimento, as moléculas em solução.
A compreensão dessa hipótese fica mais fácil acompanhando-se o modelo sugerido por Münch para a sua explicação.
A compreensão dessa hipótese fica mais fácil acompanhando-se o modelo sugerido por Münch para a sua explicação.
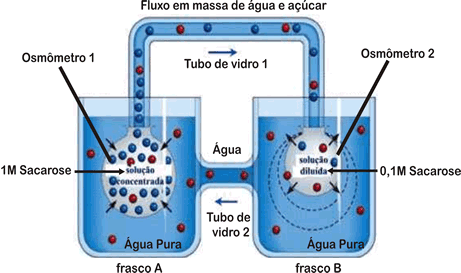
Observando a figura, conclui-se que haverá ingresso de água por osmose, do frasco A para o osmômetro 1, e do frasco B para o osmômetro 2. No entanto, como a solução do osmômetro 1 é mais concentrada, a velocidade de passagem de água do frasco A para o osmômetro 1 é maior. Assim, a água tenderá a se dirigir para o tubo de vidro 1 com velocidade, arrastando moléculas de açúcar. Como o osmômetro 2 passa a receber mais água, esta passa para o frasco B. Do frasco B, a água passa para o tubo de vidro 2, em direção ao frasco A. Podemos fazer a correspondência entre o modelo anterior e uma planta:
- Tubo de vidro 1 corresponde ao floema e o tubo de vidro 2 ao xilema;
- Osmômetro 1 corresponde a uma célula do parênquima foliar e o osmômetro 2, a uma célula da raiz;
- Frasco A representa a folha, enquanto o frasco B representa a raiz;
- As células do parênquima foliar realizam fotossíntese e produzem glicose. A concentração dessas células aumenta, o que faz com que absorvam água do xilema das nervuras. O excesso de água absorvida é deslocado para o floema, arrastando moléculas de açúcar em direção aos centros consumidores ou de reserva.
Organização dos Tecidos nas Raízes e nos Caules
Raízes e caules jovens, cortados transversalmente, mostram que são formados por uma reunião de tecidos. A disposição desses tecidos é específica em cada órgão e constitui uma estrutura interna primária típica de cada um deles. Uma estrutura secundária, mais complexa, pode ser vista quando ocorre um aumento no diâmetro do caule e da raiz.
Estrutura primária da raiz
Se acompanhássemos uma célula meristemática que terminou de surgir por mitose na extremidade de uma raiz, veríamos que ela vai se alongando, ao mesmo tempo que vai se distanciando da extremidade em decorrência do surgimento de novas células. A maior taxa de crescimento em extensão de uma raiz, ocorrerá, portanto, na região situada pouco acima da região meristemática, denominada de zona de distensão.
Após crescerem as células iniciam a sua diferenciação. Na região mais interna, por exemplo, terá início a diferenciação dos tecidos condutores, enquanto na região mais externa diferenciam-se parênquimas e tecidos de revestimento.
Após crescerem as células iniciam a sua diferenciação. Na região mais interna, por exemplo, terá início a diferenciação dos tecidos condutores, enquanto na região mais externa diferenciam-se parênquimas e tecidos de revestimento.
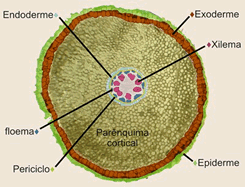
Córtex
A região mais periférica da raiz jovem diferenciam-se em epiderme, tecido formado por uma única camada de células achatadas e justapostas. Na região abaixo da epiderme, chamada córtex, diferencia-se o parênquima cortical, constituído por várias camadas de células relativamente pouco especializadas.
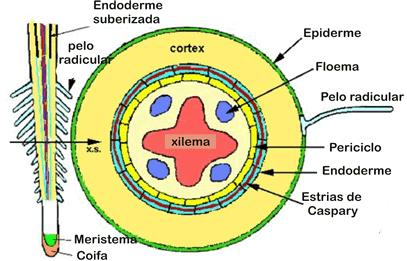
Cilindro central
A parte interna da raiz é o cilindro central, composto principalmente por elementos condutores(protoxilema e protofloema), fibras e parênquima. O cilindro central é delimitado pela endoderme, uma camada de células bem ajustadas e dotadas de reforços especiais nas paredes, as estrias de Caspary. Essas estrias são como cintas de celulose que unem firmemente as células vizinhas, vedando completamente os espaços entre elas. Assim, para penetrar no cilindro central, toda e qualquer substância tem que atravessar diretamente as células endodérmicas, uma vez que as estrias de caspary fecham os interstícios intercelulares.
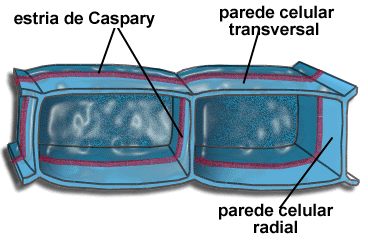
Logo abaixo da endoderme situa-se uma camada de células de paredes finas chamada periciclo, que delimita o cilindro central, onde se localizam o xilema e o floema. A maneira como os tecidos condutores se dispõem no cilindro central é um dos critérios para distinguir dicotiledôneas de monocotiledôneas.
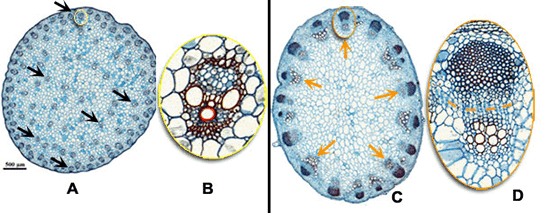
Raízes de monocotiledôneas e dicotiledôneas
Na maioria das plantas dicotiledôneas o xilema se concentra na região mias interna do cilindro central. Quando se observa um corte transversal à raiz, vê-se que o protoxilema ocupa uma área em forma de cruz ou estrela, cujas pontas encostam no periciclo. O protofloema encontra-se nos vértices formados pelos “braços” da cruz. Entre o protoxilema e o protofloema há um meristema primário chamado procâmbio. Os demais espaços dentro do cilindro central são preenchidos por parênquima.
Nas plantas monocotiledôneas, o centro da raiz é ocupado por uma medula constituída por parênquima medular e os vasos lenhosos e liberianos dispõem-se ao redor.
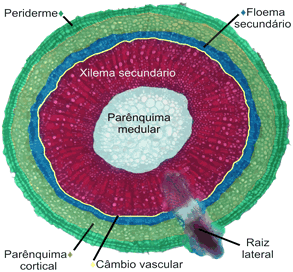
Estrutura secundária
O crescimento secundário em raízes, bem como nos caules, consiste na formação de tecidos vasculares secundários a partir do câmbio vascular e de uma periderme originada no felogênio (câmbio de casca). O câmbio vascular se inicia por divisões das células do procâmbio, que permanecem meristemáticas e estão localizadas entre o xilema e floema primários. Logo a seguir, as células do periciclo também se dividem e as células-irmãs internas, resultantes desta divisão, contribuem para formar o câmbio vascular. Um cilindro completo de câmbio da casca (felogênio), que surge na parte externa do periciclo proliferado, produz súber para o lado externo e felogênio para o lado interno. Estes três tecidos formados: súber, felogênio e feloderme constituem a periderme (RAVEN et al., 2007).
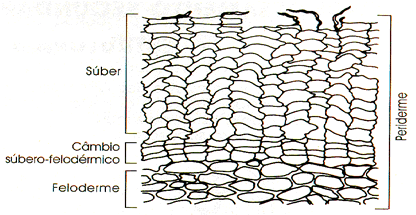
Câmbio vascular
O câmbio vascular (do latim vasculum, vaso) é assim chamado porque origina novos vasos condutores durante o crescimento secundário da raiz. O câmbio vascular forma-se a partir do procâmbio e do periciclo, que se conjugam e delimitam uma área interna do cilindro central, onde só há xilema. Ao se multiplicar ativamente, as células do câmbio vascular originam vasos xilemáticos para a região mais interna e vasos floemáticos para a região mais externa. Aos poucos a área delimitada pelo câmbio vai tornando-se cada vez mais cilíndrica.
O cambio vascular da raiz é um meristema de origem mista, primária e secundária. Isso porque tem origem tanto no procâmbio, um meristema primário, quanto do periciclo, um tecido já diferenciado que sobre desdiferenciação.
Câmbio suberógeno ou felogênio
O câmbio suberógeno, também chamado de felogênio (do grego phellos, cortiça, e genos, que gera), é um cilindro de células meristemáticas localizado na região cortical da raiz, sob a epiderme. O felogênio é um meristema secundário, uma vez que tem origem por desdiferenciação de células do parênquima cortical.
Como vimos, a atividade do felogênio produz feloderme e súber, este último um tecido morto que protege externamente raízes e caules com crescimento secundário.
Estrutura interna do caule
Como na raiz, a parte mais jovem de um caule é a que se localiza junto à extremidade, onde ocorre a multiplicação das células do meristema apical, que permite o crescimento em extensão.
Logo abaixo da zona meristemática apical as células iniciam o processo de diferenciação celular, que leva ao aparecimento dos diversos tecidos que compõem o caule.
Logo abaixo da zona meristemática apical as células iniciam o processo de diferenciação celular, que leva ao aparecimento dos diversos tecidos que compõem o caule.
Origem das gemas axilares
Nem todas as células produzidas pelo meristema apical sofrem diferenciação. À medida que o caule cresce, permanecem grupos de células meristemáticas sob a epiderme, pouco acima do ponto de inserção das folhas. Esses grupos de células formam, nas axilas das folhas, protuberâncias chamadas gemas axilares ou laterais.
Estrutura primária
Feixes líbero-lenhosos
Caules que não cresceram, em espessura apresentam estrutura primária, caracterizada pela presença de feixes líbero-lenhosos localizados entre as células do parênquima que preenchem seu interior.
Cada feixe libero-lenhoso possui elementos do líber (floema) voltados para fora e elementos do lenho (xilema) voltados para dentro.
Nas plantas monocotiledôneas, que geralmente na apresentam crescimento secundário, os feixes condutores são distribuídos de maneira difusa no interior do caule. Já nas dicotiledôneas os feixes líbero-lenhosos distribuem-se regularmente, formando um cilindro.
Câmbio fascicular
Nos feixes libero-lenhosos das dicotiledôneas, o floema está voltado para o exterior do caule e o xilema para o interior. Entre o floema e o xilema de um feixe há um tecido meristemático: o câmbio vascular e o câmbio suberógeno ou felogênio.
Câmbio vascular
O Câmbio vascular do caule forma-se a partir do câmbio fascicular e do câmbio interfascicular, este último um tecido meristemátco secundário, resultante da dedisferenciação de células parenquimáticas localizadas entre os feixes libero-lenhosos. O câmbio fascicular passa a delimitar, assim, uma área interna do caule onde só há xilema e parênquima. Como na raiz, as células do câmbio vascular originam vasos xilemáticos para a região mais interna e vasos floemáticos para a região mais externa. Aos poucos, a área delimitada pelo câmbio vai se tornando cada vez mais cilíndrica.
O câmbio vascular o caule, como o da raiz, também é um meristema de origem mista, primária e secundária. Isso porque tem origem tanto do cambio fascicular, um meristema primário, quanto do câmbio interfascicular, um meristema que surgiu da desdiferenciação de células parenquimáticas.
Câmbio suberógeno ou felogênio
A atividade do câmbio vascular faz com que o caule vá progressivamente aumentando de espessura. Para acompanhar esse crescimento em diâmetro, célula do parênquima cortical sofrem desdiferenciação e originam um cilindro de meristema secundário, o felogênio. Como vimos, a atividade do felogênio produz feloderme para o interior e súber para o exterior, formando a periderme, que passa a revestir o caule.
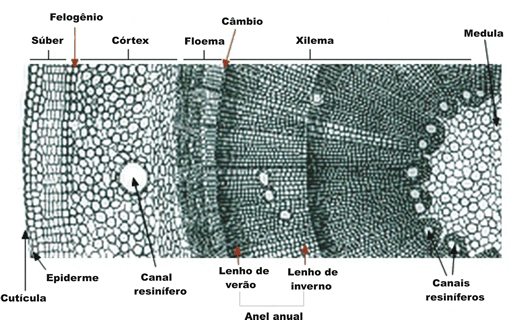
Anéis anuais
Nas regiões de clima temperado, a atividade do câmbio varia no decorrer do ano. A atividade cambial é muito intensa durante a primavera e o verão, diminuindo progressivamente no outono até cessar por completo no inverno. No fim do verão, quando está encerrando mais um ciclo de atividade, o câmbio produz vasos lenhosos de paredes grossas e lúmen estreito, que constituem o lenho estival. Na primavera, ao retomar o seu funcionamento depois do repouso invernal, o câmbio produz vasos lenhosos de paredes delgadas e lúmen grande, que constituem o lenho primaveril.
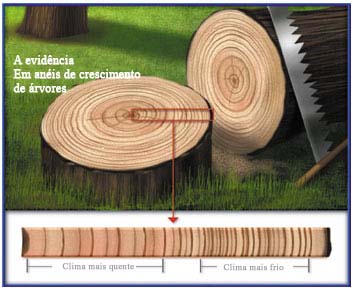
Troncos de árvores que vivem em regiões temperadas apresentam, portanto, anéis de lenho primaveril. Quando esses troncos são observados em corte transversal, esses anéis são facilmente identificáveis e o número de pares de anéis corresponde à idade da árvore.
Crescimento e desenvolvimento
O crescimento de uma planta começa a partir da germinação da semente. A hidratação da semente, por exemplo, ativa o embrião. As reservas contidas no endosperma ou nos cotilédones são hidrolisadas por ação enzimática. As células embrionárias recebem os nutrientes necessários, o metabolismo aumenta e são iniciadas as divisões celulares que conduzirão ao crescimento.
A radícula é a primeira estrutura a imergir; a seguir, exterioriza-se o caulículo e a plântula inicia um longo processo que culminará no vegetal adulto.
A radícula é a primeira estrutura a imergir; a seguir, exterioriza-se o caulículo e a plântula inicia um longo processo que culminará no vegetal adulto.
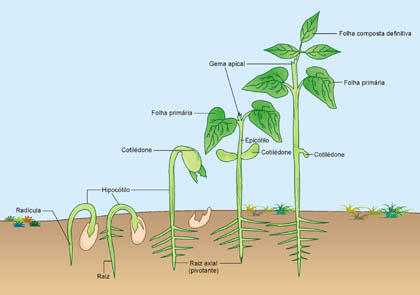
Qual a diferença entre crescimento e desenvolvimento?
Esses dois termos são frequentemente utilizados como sinônimos. No entanto, há uma diferença entre eles:
- O crescimento corresponde a um crescimento irreversível no tamanho de um vegetal, e se dá a partir do acréscimo de células resultantes das divisões mitóticas, além do tamanho individual de cada célula. De modo geral, o crescimento também envolve aumento do volume e da massa do vegetal. O crescimento envolve parâmetros quantitativos mensuráveis (tamanho, massa e volume).
- O desenvolvimento consiste no surgimento dos diferentes tipos celulares e dos diversos tecidos componentes dos órgão vegetais. É certamente um fenômeno relacionado ao processo de diferenciação celular. O desenvolvimento envolve aspectos quantitativos, relacionados ao aumento da complexidade do vegetal.
A ocorrência desses dois processos é simultânea. Um vegetal cresce e se desenvolve ao mesmo tempo.
O meristema
Todos os tipos de células que compõe uma planta tiveram origem a partir de tecidos meristemáticos, formados por células que têm uma parede primária fina, pequenos vacúolos e grande capacidade de realiza mitose.
As células dos tecidos diferenciados, ainda que não tenham morrido durante a diferenciação (como o súber, o xilema, por exemplo), perdem a capacidade de se multiplicar por mitose. As células meristemáticas se multiplicam e se diferenciam, originando os diversos tecidos permanentes da planta, cujas células não mais se dividem.
Meristema primários
Em certos locais das plantas, como nos ápices da raiz e do caule, existem tecidos meristemáticos que descendem diretamente das primeiras células embrionários, presentes na semente. Esses são os meristemas primários.
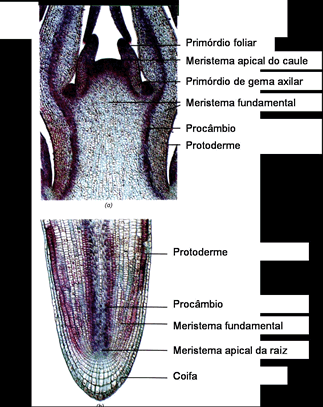
O caule cresce em comprimento graças à atividade de um meristema primário presente em seu ápice, o meristema apical caulinar. Já o meristema responsável pelo crescimento em comprimento da raiz não é terminal, mas está protegido sob um capuz celular chamado de coifa. Por isso é denominadomeristema subapical radicular. | 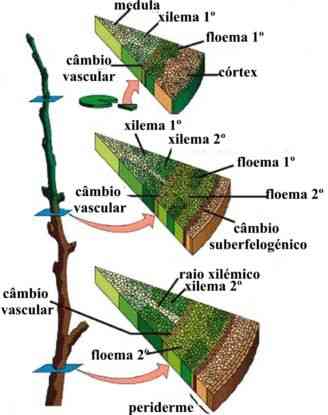 |
Meristemas secundários
Meristemas secundários são os que surgem a partir de células diferenciadas, geralmente parenquimáticas, que readquirem a capacidade mitótica, fenômeno que os botânicos denominam desdiferenciação. O felogênio que constitui a periderme, por exemplo, é um exemplo de meristema secundário, que surge pela desdiferenciação de células do parênquima localizadas sob a epiderme. A multiplicação das células do felogênio origina a feloderme e o súber que compõem a periderme.
http://www.portalsaofrancisco.com.br/alfa/esclerenquima/esclerenquima-1.php
http://www.sobiologia.com.br/conteudos/Morfofisiologia_vegetal/morfovegetal26.php
Exercícios sobre tecidos vegetais
1) Os meristemas vegetais, quando examinados ao microscópio, apresentam abundância de:
a) reservas
b) figuras de mitose
c) figuras de meiose
d) tecido lenhoso
e) tecido vascular
2) (UFCE) Nas células meristemáticas, é comum a observação de:
a) grandes vacúolos
b) citoplasma parietal
c) figuras mitóticas
d) divisão meiótica
e) núcleo periférico
3) (UFBA) Qual dos seguintes conjuntos de características é comum a todos os tecidos de sustentação dos vegetais?
a) células mortas, localização periférica e presença de lignina;
b) células em atividade, localização interna e parede reforçada com substâncias diversas;
c) células mortas ou em atividade, localização variada e parede reforçada com substâncias diversas;
d) células alongadas, localização periférica e presença de lignina ou de celulose;
e) células alongadas, mortas, localização interna e parede reforçada com substâncias diversas.
4) (UFSE) A conhecida cortiça, de tão larga aplicação na fabricação de rolhas, é retirada de árvores que apresentam abundância do tecido denominado:
a) súber
b) lenho
c) colênquima
d) esclerênquima
e) líber
5) Quais são as estruturas vegetais relacionadas com as trocas gasosas nos vegetais?
6) (PUC) O câmbio e o felogênio são duas formações vegetais constituídas por tecido:
a) meristemático de crescimento;
b) meristemático, de crescimento em comprimento, existente na raiz;
c) diferenciado, de crescimento, existente no caule e raiz;
d) diferenciado para condução, existente nas angiospermas;
e) diferenciado para promover absorção de água, existente nas raízes.
7) (UFRJ 2006) Tal como acontece com os animais, os vegetais superiores também apresentam células com uma organização estrutural formando tecidos. Existe uma certa analogia entre alguns tecidos vegetais e determinados tecidos animais. Esta analogia existe entre:
a) o esclerênquima encontrado nos vegetais e tecido ósseo dos animais.
b) o tecido suberoso dos vegetais e o tecido sanguíneo dos animais.
c) os vasos liberianos dos vegetais e o tecido ósseo dos animais.
d) os canais laticíferos dos vegetais e a epiderme dos animais.
e) o colênquima dos vegetais e o tecido muscular liso dos animais.
8) (UFV) Em relação aos tecidos vegetais:
a) qual a função dos meristemas primários e onde se localizam?
b) qual a função dos meristemas secundários?
c) dê o nome do tecido localizado nas folhas e nos caules jovens, caracterizados por células ricamente clorofiladas com função fotossintética.
9) (FUVEST) O que são meristemas e quais seus principais tipos?
10) (FUVEST) Cite dois tipos de parênquimas e suas funções.
11) (FURG) Considerando os tecidos vegetais, relacione a coluna 1 com a coluna 2.
Coluna 1
I - Tecido que garante a flutuação de plantas aquáticas.
II - Tecido que garante a sobrevivência de plantas em ambientes secos como desertos.
III - Tecido responsável pelo crescimento em comprimento de caules e raízes.
IV - Tecido responsável pelo crescimento em espessura dos caules.
V - Tecido responsável pela fotossíntese.
VI - Tecido de características mecânicas que serve para a sustentação da planta.
Coluna 2
( ) Câmbio
( ) Meristema apical
( ) Esclerênquima
( ) Parênquima clorofiliano
( ) Parênquima aerífero
( ) Parênquima aqüífero
Assinale a alternativa com a seqüência correta
a) I, II, III, V, VI e IV.
b) III, I, II, IV, V e VI.
c) III, IV, VI, I, II e V.
d) IV, VI, III, V, I e II.
e) IV, III, VI, V, I e II.
12) As células epidérmicas das raízes (pêlos absorventes) absorvem água do solo, normalmente, quando:
a) a concentração de sais das células for menor que a concentração de sais do solo.
b) a concentração de sais das células for maior que a concentração de sais do solo.
c) a concentração de sais das células for igual à concentração de sais do solo.
d) a concentração de água das células for maior que a concentração de água do solo.
e) a concentração de água das células for igual à concentração de água do solo.
13) (Cesgranrio - RJ) Nas plantas, as estruturas especializadas para as trocas de gases entre a planta e o exterior são:
a) os estômatos e as lenticelas.
b) o floema e os estômatos.
c) as lenticelas e o colênquima.
d) o esclerênquima e o xilema.
e) o súber e o câmbio.
14) (Fuvest-SP) O xilema ou lenho é responsável:
a) pela absorção de água e sais minerais.
b) pela condução de substâncias orgânicas liberadas pelo órgão de reserva.
c) pelo transporte e pela distribuição de água e nutrientes minerais.
d) pelo transporte e pela distribuição de alimentos orgânicos.
e) pelo transporte de água e alimentos orgânicos sintetizados na folha.
15) (MACK-SP) Alguns tecidos vegetais são constituídos por células mortas, não como uma medida casual, mas como ponto final de um processo de diferenciação celular tão importante como qualquer outra função desempenhada por qualquer outra célula viva. São exemplos de tecidos vegetais constituídos por células mortas:
a) colênquima e parênquima paliçádico.
b) súber e parênquima paliçádico.
c) esclerênquima e colênquima.
d) súber e colênquima.
e) súber e esclerênquima.
16) Quais os principais componentes do xilema e qual a sua função?
17) (UFV) Embora não sejam essenciais como nutriente, as fibras têm papel importante na dieta. Elas estimulam os movimentos peristálticos, reduzindo o tempo de contato de substâncias potencialmente prejudiciais no intestino, evitando assim muitos distúrbios como diverticulose e prisão de ventre. A sua deficiência na dieta pode causar também problemas mais graves como doenças coronarianas, câncer no intestino e obesidade. Em relação às fibras, resolva os itens abaixo.
a) Cite um tecido vegetal rico em fibras.
b) Além da alimentação, cite uma importância econômica de utilização dessas fibras.
18) (UFU-MG) Comparando-se os tecidos vegetais pede-se uma semelhança e uma diferença entre cada um dos pares abaixo:
a) Colênquima e esclerênquima.
b) Meristema primário e meristema secundário.
19) (FUVEST) Muitas fibras do esclerênquima são usadas industrialmente como matéria-prima para a fabricação do cânhamo, da juta e do linho.
a) Cite duas características do esclerênquima.
b) Identifique a principal função desse tecido vegetal.
20) (FUVEST-SP) O esquema representa um corte transversal de um tronco de árvore.
 a) Em quais tecidos indicados espera-se encontrar células em divisão?
a) Em quais tecidos indicados espera-se encontrar células em divisão?
b) Em qual dos tecidos indicados espera-se encontrar seiva com maior concentração de substâncias orgânicas?
21) (UFPR) Tal como sucede com os animais, também as plantas desenvolvidas apresentam as suas células com uma organização estrutural formando tecidos. Os tecidos vegetais se distribuem em dois grandes grupos: tecidos de formação e tecidos permanentes. Com relação aos tecidos vegetais, assinale as proposições CORRETAS.
(01) Os meristemas e a epiderme são exemplos de tecidos de formação.
(02) O xilema e o colênquima são tecidos permanentes.
(04) Os meristemas são tecidos embrionários dos quais resultam todos os demais tecidos vegetais.
(08) Os parênquimas, quando dotados de células ricamente clorofiladas, são tecidos de síntese.
(16) Os tecidos de arejamento se destinam às trocas gasosas e de sais minerais entre a planta e o meio ambiente, sendo o floema um de seus principais exemplos.
(32) As bolsas secretoras, presentes em nectários, juntamente com os canais laticíferos, existentes nas seringueiras, são exemplos de tecidos de secreção.
Soma ( )
22) (UFPR) Assinale as proposições que apresentam CORRETAMENTE os tipos de raízes, suas características e exemplos de vegetais que as possuem.
(01) Raízes fasciculadas possuem um eixo central mais desenvolvido e estão presentes nos vegetais chamados dicotiledôneas, como o milho e a grama.
(02) As raízes respiratórias, presentes nos mangues, promovem uma maior absorção de oxigênio.
(04) Raízes tabulares aumentam a estabilidade de árvores frondosas, como as figueiras.
(08) As raízes sugadoras dos vegetais parasitas, como o cipó-chumbo, penetram nos caules de plantas hospedeiras, sugando a seiva de que necessitam.
(16) A raiz tuberosa, presente na cenoura e beterraba, acumula substâncias nutritivas.
Soma ( )
23) (UNESP) São exemplos de tecidos de sustentação, condução e proteção, respectivamente:
a) súber - traqueídeos - esclerênquima.
b) epiderme - esclerênquima - súber.
c) súber - colênquima - fibras.
d) esclerênquima - traqueídeos - súber.
e) colênquima - xilema - traqueídeos.
24) (UERJ) Até cerca de 405 milhões de anos atrás, parece que a vida esteve limitada à água. A existência terrestre trouxe consigo sérios problemas como, por exemplo, o risco de dessecamento. Características que permitissem aos vegetais a redução de perda d'água em suas partes aéreas foram selecionadas positivamente pelo ambiente por facilitar a adaptação.
A economia de água é permitida pelo seguinte tecido vegetal:
a) súber
b) floema
c) colênquima
d) parênquima de assimilação.
25) (UFRRJ) Sobre o esquema a seguir são feitas algumas afirmativas:

/Softcom/SV%20v3/Dados/Users/adilson/Arquivos/Figuras/6533)
Sobre as afirmativas, pode-se concluir que apenas
I) O esquema representa o tecido vegetal de sustentação.
II) Neste sistema movimenta-se uma solução orgânica onde predominam açúcares solúveis.
III) Este tecido está presente em todos os vegetais terrestres.
IV) A movimentação de solução orgânica neste sistema faz-se da região mais concentrada para a menos concentrada.
a) II e III estão corretas.
b) II e IV estão corretas.
c) I e IV estão corretas.
d) I e II estão corretas.
e) I e III estão corretas.
Gabarito:
http://www.sobiologia.com.br/conteudos/Morfofisiologia_vegetal/morfovegetal26.php
Exercícios sobre tecidos vegetais
1) Os meristemas vegetais, quando examinados ao microscópio, apresentam abundância de:
a) reservas
b) figuras de mitose
c) figuras de meiose
d) tecido lenhoso
e) tecido vascular
2) (UFCE) Nas células meristemáticas, é comum a observação de:
a) grandes vacúolos
b) citoplasma parietal
c) figuras mitóticas
d) divisão meiótica
e) núcleo periférico
3) (UFBA) Qual dos seguintes conjuntos de características é comum a todos os tecidos de sustentação dos vegetais?
a) células mortas, localização periférica e presença de lignina;
b) células em atividade, localização interna e parede reforçada com substâncias diversas;
c) células mortas ou em atividade, localização variada e parede reforçada com substâncias diversas;
d) células alongadas, localização periférica e presença de lignina ou de celulose;
e) células alongadas, mortas, localização interna e parede reforçada com substâncias diversas.
4) (UFSE) A conhecida cortiça, de tão larga aplicação na fabricação de rolhas, é retirada de árvores que apresentam abundância do tecido denominado:
a) súber
b) lenho
c) colênquima
d) esclerênquima
e) líber
5) Quais são as estruturas vegetais relacionadas com as trocas gasosas nos vegetais?
6) (PUC) O câmbio e o felogênio são duas formações vegetais constituídas por tecido:
a) meristemático de crescimento;
b) meristemático, de crescimento em comprimento, existente na raiz;
c) diferenciado, de crescimento, existente no caule e raiz;
d) diferenciado para condução, existente nas angiospermas;
e) diferenciado para promover absorção de água, existente nas raízes.
7) (UFRJ 2006) Tal como acontece com os animais, os vegetais superiores também apresentam células com uma organização estrutural formando tecidos. Existe uma certa analogia entre alguns tecidos vegetais e determinados tecidos animais. Esta analogia existe entre:
a) o esclerênquima encontrado nos vegetais e tecido ósseo dos animais.
b) o tecido suberoso dos vegetais e o tecido sanguíneo dos animais.
c) os vasos liberianos dos vegetais e o tecido ósseo dos animais.
d) os canais laticíferos dos vegetais e a epiderme dos animais.
e) o colênquima dos vegetais e o tecido muscular liso dos animais.
8) (UFV) Em relação aos tecidos vegetais:
a) qual a função dos meristemas primários e onde se localizam?
b) qual a função dos meristemas secundários?
c) dê o nome do tecido localizado nas folhas e nos caules jovens, caracterizados por células ricamente clorofiladas com função fotossintética.
9) (FUVEST) O que são meristemas e quais seus principais tipos?
11) (FURG) Considerando os tecidos vegetais, relacione a coluna 1 com a coluna 2.
Coluna 1
I - Tecido que garante a flutuação de plantas aquáticas.
II - Tecido que garante a sobrevivência de plantas em ambientes secos como desertos.
III - Tecido responsável pelo crescimento em comprimento de caules e raízes.
IV - Tecido responsável pelo crescimento em espessura dos caules.
V - Tecido responsável pela fotossíntese.
VI - Tecido de características mecânicas que serve para a sustentação da planta.
Coluna 2
( ) Câmbio
( ) Meristema apical
( ) Esclerênquima
( ) Parênquima clorofiliano
( ) Parênquima aerífero
( ) Parênquima aqüífero
Assinale a alternativa com a seqüência correta
a) I, II, III, V, VI e IV.
b) III, I, II, IV, V e VI.
c) III, IV, VI, I, II e V.
d) IV, VI, III, V, I e II.
e) IV, III, VI, V, I e II.
12) As células epidérmicas das raízes (pêlos absorventes) absorvem água do solo, normalmente, quando:
a) a concentração de sais das células for menor que a concentração de sais do solo.
b) a concentração de sais das células for maior que a concentração de sais do solo.
c) a concentração de sais das células for igual à concentração de sais do solo.
d) a concentração de água das células for maior que a concentração de água do solo.
e) a concentração de água das células for igual à concentração de água do solo.
13) (Cesgranrio - RJ) Nas plantas, as estruturas especializadas para as trocas de gases entre a planta e o exterior são:
a) os estômatos e as lenticelas.
b) o floema e os estômatos.
c) as lenticelas e o colênquima.
d) o esclerênquima e o xilema.
e) o súber e o câmbio.
14) (Fuvest-SP) O xilema ou lenho é responsável:
a) pela absorção de água e sais minerais.
b) pela condução de substâncias orgânicas liberadas pelo órgão de reserva.
c) pelo transporte e pela distribuição de água e nutrientes minerais.
d) pelo transporte e pela distribuição de alimentos orgânicos.
e) pelo transporte de água e alimentos orgânicos sintetizados na folha.
15) (MACK-SP) Alguns tecidos vegetais são constituídos por células mortas, não como uma medida casual, mas como ponto final de um processo de diferenciação celular tão importante como qualquer outra função desempenhada por qualquer outra célula viva. São exemplos de tecidos vegetais constituídos por células mortas:
a) colênquima e parênquima paliçádico.
b) súber e parênquima paliçádico.
c) esclerênquima e colênquima.
d) súber e colênquima.
e) súber e esclerênquima.
16) Quais os principais componentes do xilema e qual a sua função?
17) (UFV) Embora não sejam essenciais como nutriente, as fibras têm papel importante na dieta. Elas estimulam os movimentos peristálticos, reduzindo o tempo de contato de substâncias potencialmente prejudiciais no intestino, evitando assim muitos distúrbios como diverticulose e prisão de ventre. A sua deficiência na dieta pode causar também problemas mais graves como doenças coronarianas, câncer no intestino e obesidade. Em relação às fibras, resolva os itens abaixo.
a) Cite um tecido vegetal rico em fibras.
b) Além da alimentação, cite uma importância econômica de utilização dessas fibras.
18) (UFU-MG) Comparando-se os tecidos vegetais pede-se uma semelhança e uma diferença entre cada um dos pares abaixo:
a) Colênquima e esclerênquima.
b) Meristema primário e meristema secundário.
19) (FUVEST) Muitas fibras do esclerênquima são usadas industrialmente como matéria-prima para a fabricação do cânhamo, da juta e do linho.
a) Cite duas características do esclerênquima.
b) Identifique a principal função desse tecido vegetal.
20) (FUVEST-SP) O esquema representa um corte transversal de um tronco de árvore.
b) Em qual dos tecidos indicados espera-se encontrar seiva com maior concentração de substâncias orgânicas?
21) (UFPR) Tal como sucede com os animais, também as plantas desenvolvidas apresentam as suas células com uma organização estrutural formando tecidos. Os tecidos vegetais se distribuem em dois grandes grupos: tecidos de formação e tecidos permanentes. Com relação aos tecidos vegetais, assinale as proposições CORRETAS.
(01) Os meristemas e a epiderme são exemplos de tecidos de formação.
(02) O xilema e o colênquima são tecidos permanentes.
(04) Os meristemas são tecidos embrionários dos quais resultam todos os demais tecidos vegetais.
(08) Os parênquimas, quando dotados de células ricamente clorofiladas, são tecidos de síntese.
(16) Os tecidos de arejamento se destinam às trocas gasosas e de sais minerais entre a planta e o meio ambiente, sendo o floema um de seus principais exemplos.
(32) As bolsas secretoras, presentes em nectários, juntamente com os canais laticíferos, existentes nas seringueiras, são exemplos de tecidos de secreção.
Soma ( )
22) (UFPR) Assinale as proposições que apresentam CORRETAMENTE os tipos de raízes, suas características e exemplos de vegetais que as possuem.
(01) Raízes fasciculadas possuem um eixo central mais desenvolvido e estão presentes nos vegetais chamados dicotiledôneas, como o milho e a grama.
(02) As raízes respiratórias, presentes nos mangues, promovem uma maior absorção de oxigênio.
(04) Raízes tabulares aumentam a estabilidade de árvores frondosas, como as figueiras.
(08) As raízes sugadoras dos vegetais parasitas, como o cipó-chumbo, penetram nos caules de plantas hospedeiras, sugando a seiva de que necessitam.
(16) A raiz tuberosa, presente na cenoura e beterraba, acumula substâncias nutritivas.
Soma ( )
23) (UNESP) São exemplos de tecidos de sustentação, condução e proteção, respectivamente:
a) súber - traqueídeos - esclerênquima.
b) epiderme - esclerênquima - súber.
c) súber - colênquima - fibras.
d) esclerênquima - traqueídeos - súber.
e) colênquima - xilema - traqueídeos.
24) (UERJ) Até cerca de 405 milhões de anos atrás, parece que a vida esteve limitada à água. A existência terrestre trouxe consigo sérios problemas como, por exemplo, o risco de dessecamento. Características que permitissem aos vegetais a redução de perda d'água em suas partes aéreas foram selecionadas positivamente pelo ambiente por facilitar a adaptação.
A economia de água é permitida pelo seguinte tecido vegetal:
a) súber
b) floema
c) colênquima
d) parênquima de assimilação.
25) (UFRRJ) Sobre o esquema a seguir são feitas algumas afirmativas:

Sobre as afirmativas, pode-se concluir que apenas
I) O esquema representa o tecido vegetal de sustentação.
II) Neste sistema movimenta-se uma solução orgânica onde predominam açúcares solúveis.
III) Este tecido está presente em todos os vegetais terrestres.
IV) A movimentação de solução orgânica neste sistema faz-se da região mais concentrada para a menos concentrada.
a) II e III estão corretas.
b) II e IV estão corretas.
c) I e IV estão corretas.
d) I e II estão corretas.
e) I e III estão corretas.
Gabarito:
1) B 2) C 3) C 4) A 5) Epiderme – estômatos Súber – lenticelas 6) A 7) A 8) a) Localizados no ápice e nas gemas laterais do caule e na região subapical da raiz. Os meristemas primários atuam no crescimento longitudinal ou em comprimento da raiz e do caule. b) Crescimento do vegetal em espessura (tranversal). c) Parênquima clorofiliano ou assimilador. 9) São tecidos formados por células com alta capacidade de divisão celular, ou seja, dotados de alto poder proliferativo. Há dois tipos de tecidos meristemáticas: primários e secundários. 10) Poderiam ser citados os seguintes tipos de parênquimas: clorofiliano cuja função é a fotossíntese; amilífero que possui a função de armazenar o amido; aqüífero cuja função é armazenar água em vegetais xerófitos; aerífero que possui a função de armazenar ar nas plantas aquáticas. 11) E 12) B 13) A 14) C 15) E 16) O xilema ou lenho é formado por: vasos lenhosos, parênquima lenhoso e fibras de sustentação. Os vasos lenhosos são formados por células mortas lignificas. O parênquima lenhoso é formado por células vivas, com função de preenchimento. Os elementos de sustentação são formados por fibras do esclerênquima. O xilema tem a função de transportar a seiva mineral ou bruta das raízes para as folhas. 17) a) Esclerênquima. b) Produção do linho. 18) a) Colênquima e esclerênquima são tecidos de sustentação, formado por células que apresentam paredes espessas. Diferem pelo fato de o colênquima ser formado por células vivas cuja parede celular tem reforço de celulose, enquanto o esclerênquima é formado por células mortas e paredes com reforço de lignina.
b) Meristemas primários e secundários são formados por células indiferenciadas, com grande capacidade de divisão celular, sendo responsáveis pelo crescimento dos vegetais. Diferem pelo fato de os meristemas primários determinarem crescimento longitudinal ou em comprimento e os meristemas secundários, o crescimento em espessura ou transversal da raiz e do caule. 19) a) O esclerênquima é composto de células mortas, alongadas com paredes espessas e resistentes. Devido à presença de lignina.
b) A função do esclerênquima é a sustentação dos vegetais.
20) a) Células em divisão serão encontradas nos meristemas, que são câmbio e o felogênio. Ambos são tecidos meristemáticos secundários.
b) O floema transporta a seiva elaborada, rica em substâncias orgânicas, principalmente açúcares.
b) Meristemas primários e secundários são formados por células indiferenciadas, com grande capacidade de divisão celular, sendo responsáveis pelo crescimento dos vegetais. Diferem pelo fato de os meristemas primários determinarem crescimento longitudinal ou em comprimento e os meristemas secundários, o crescimento em espessura ou transversal da raiz e do caule. 19) a) O esclerênquima é composto de células mortas, alongadas com paredes espessas e resistentes. Devido à presença de lignina.
b) A função do esclerênquima é a sustentação dos vegetais.
20) a) Células em divisão serão encontradas nos meristemas, que são câmbio e o felogênio. Ambos são tecidos meristemáticos secundários.
b) O floema transporta a seiva elaborada, rica em substâncias orgânicas, principalmente açúcares.
21) 02 + 04 + 08 + 32 = 46 22) 02 + 04 + 08 + 16 = 30 23) D 24) A 25) B
Nenhum comentário:
Postar um comentário